Neurobiology of interval timing published: 10th July 2025
What, then, is time? If no one asks of me, I know; if I wish to explain to him who asks, I know not.
- Saint Augustine
For centuries, humans have thought about time, its nature and how we process times in our minds. Time perception is part of the human experience yet the complete neural basis of time perception is still unknown. One of the most fundamental variables that organisms have to quantify in order to survive is temporality or time. Our brains have to develop a sense of time to do activities like talk, sing, run, perform activities in a group, drive, etc. Humans unfortunately do not posses a sensory organ to detect time like we do for audio and visual sensory input yet we have a sense of time. Time thus is not a material object of the world for which we could have a sensory organ to sense it. On a scale from microseconds on one side and circadian rhythm on the other, temporal information guides our behaviour. A 12 order magnitude is covered by this scale.
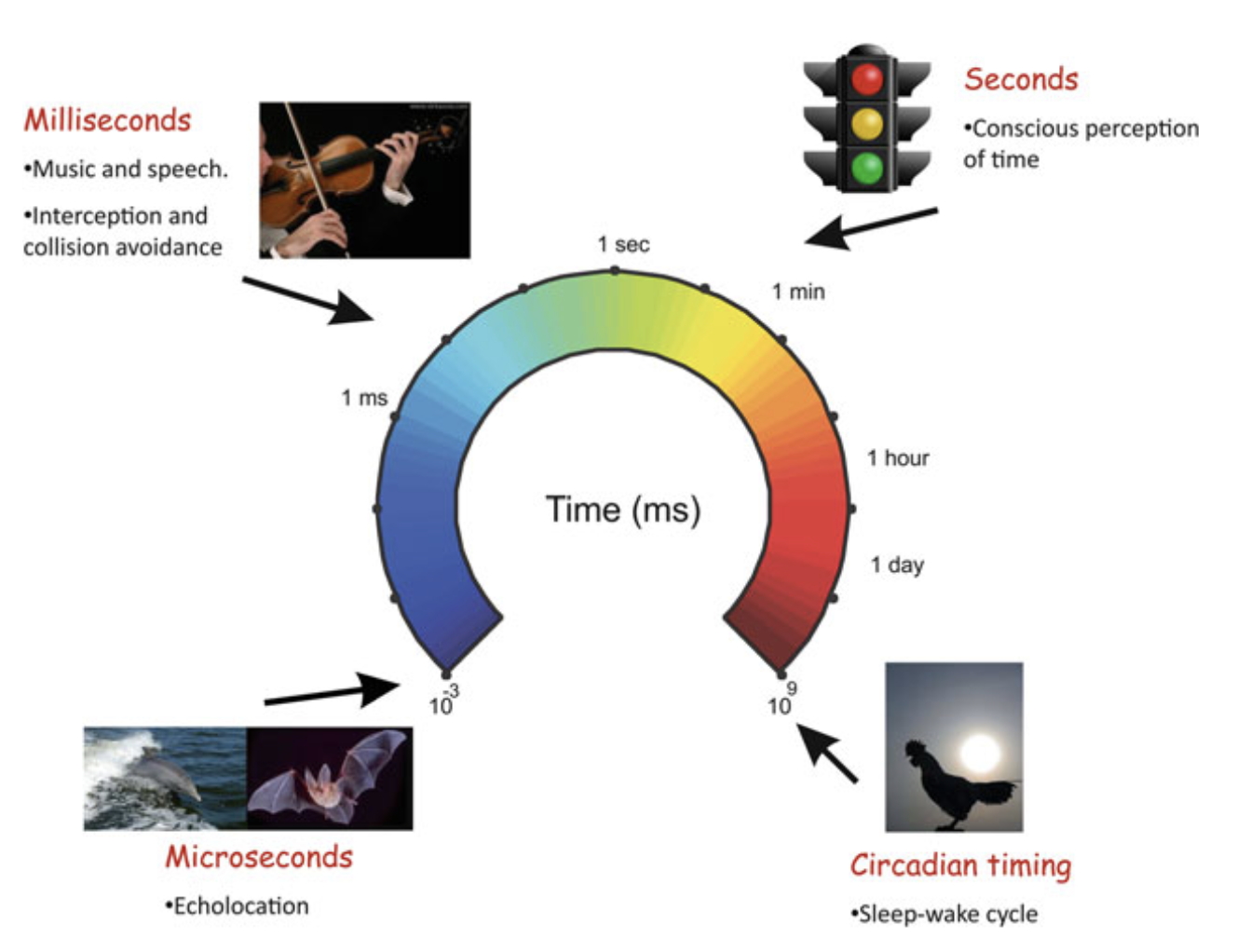
Microsecond Scale
Information processing in the microseconds is needed for binaural hearing and echolocation. For binaural hearing the microsecond scale is used by the auditory system to determine the differences in time in the action potentials for the two ears called as interaural delay to determine the spatial origin of the sound. The sound will arrive slightly earlier for the near ear usually in the order of 600-700μs because of the natural interaural time difference (ITD).
Milliseconds Range
Intercal timing ranging from 200-1000ms involves a broad spectrum of activities ranging from collision avoidance, object interception, speech perception, motion processing, execution and processing music, coordinating fine movements, etc. The temporal processing in this range is quite sophisticated yet poorly understood on how the neural processing works to make it happen.
Seconds to Minute Scale
Temporal processing in this range depends on conscious and cognitive control and is involved in foraging, decision making, sequential motor performance, arithmetic, associative learning, etc. Timing in this scale forms the contextual framework through which our behaviour is mapped onto the external world.
Circadian Rhythms
The biological timing that organises the daily environmental
oscillations every 24 hours is known as the circadian clock. The two
major functions of this clock are:
- Prepare your body for what's coming next based on the time of day.
It optimises the temporal manifestations of different biological
activities during the day by anticipating of recurring fluctuations in
the environment.
- Prevent biological conflicts by timing different processes to not
interfere with each other. Eg. release growth hormones only while
sleeping and not during the day when you need energy for activities.
The circadian rhythm in mammals is governed by a complex network of
cellular molecular oscillators distributed
throughout the brain and peripheral tissues. The master clock is in
the brain in this region known as
hypothalamic suprachiasmatic nuclei (SCN).
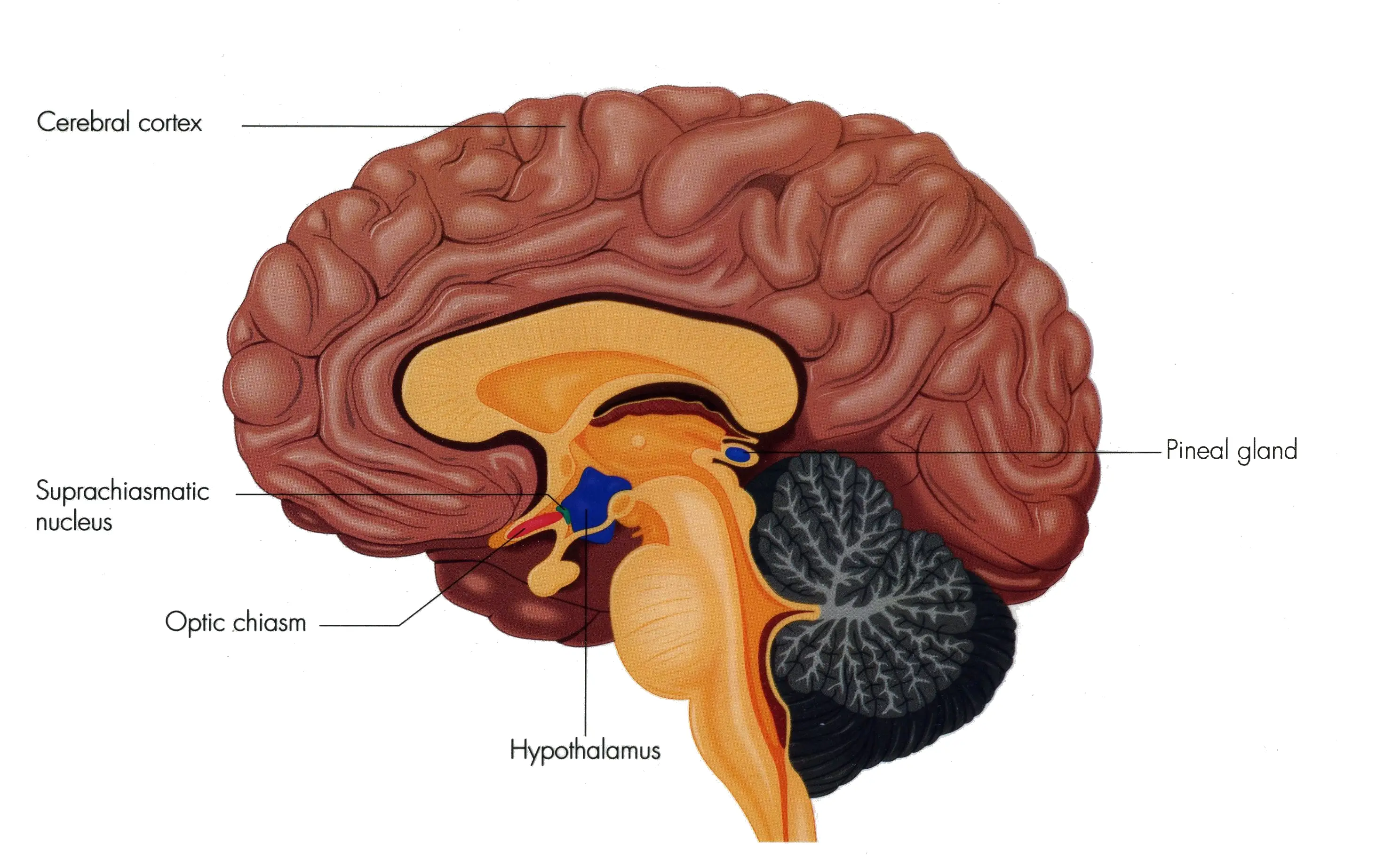
It synchronises the internal clock with the external light-dark cycle of the day. The internal circadian clock in mammals possesses a rhythm with an approximate 24 hour period and the synchronising signal is light. The SCN acts as a relay between the external light-dark cycle and the internal timing system. So, the peripheral clocks are governed directly by the SCN that controls the activities like feeding, body temperature, hormone release, etc.
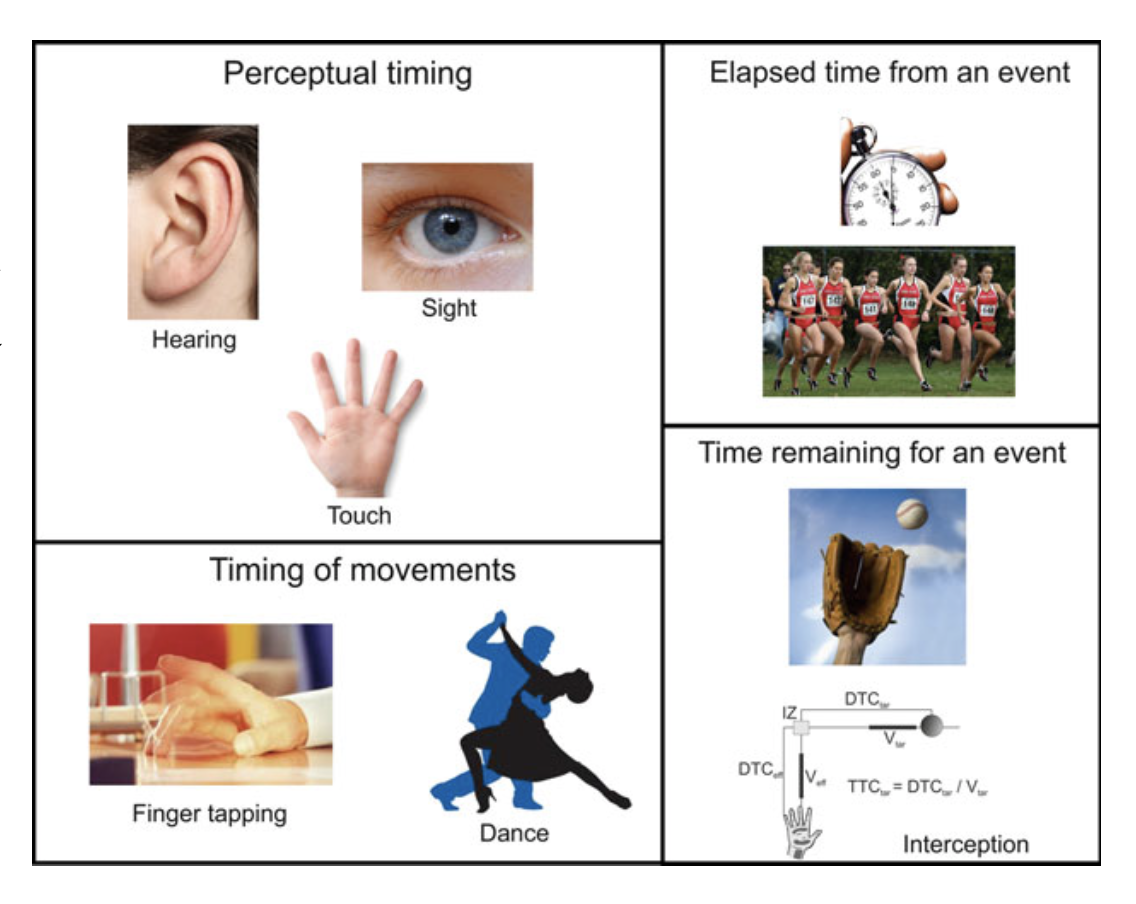
Different timing mechanism for different timing behaviours?
The word timing can have the connotation
of either how long an event lasts or
when an event occurs. So, the internal
clocks should have the ability to encode the elapsed time from a
stimulus or process such as the time between the notes in a song,
along with the capacity to measure remaining time for an action where
the system should select a precise moment for doing something for an
accurate result like when a batsman hits a cricket ball.
Our brains need to compute time from different modalities of sensory
stimulus or when involved in rhythmic activities like music and
dancing or when marking the elapsed time from an event or as the time
remaining for an action. So, some key elements of temporal processing
include the time scale being quantified, the modality of the stimuli
guiding the time perception, whether the time is being measured for a
movement or perceptual decision, whether the task involves single or
multiple intervals, whether the timing generated is external or
internal.
Cognitive model of time
Research from experimental psychology, clinical neuropsychology, and
neuroimaging have produced an extensive literature on the cognitive
mechanism of temporal processing. Cognitive models distinguish time
perception with two fundamental views:
prospective and
retrospective.
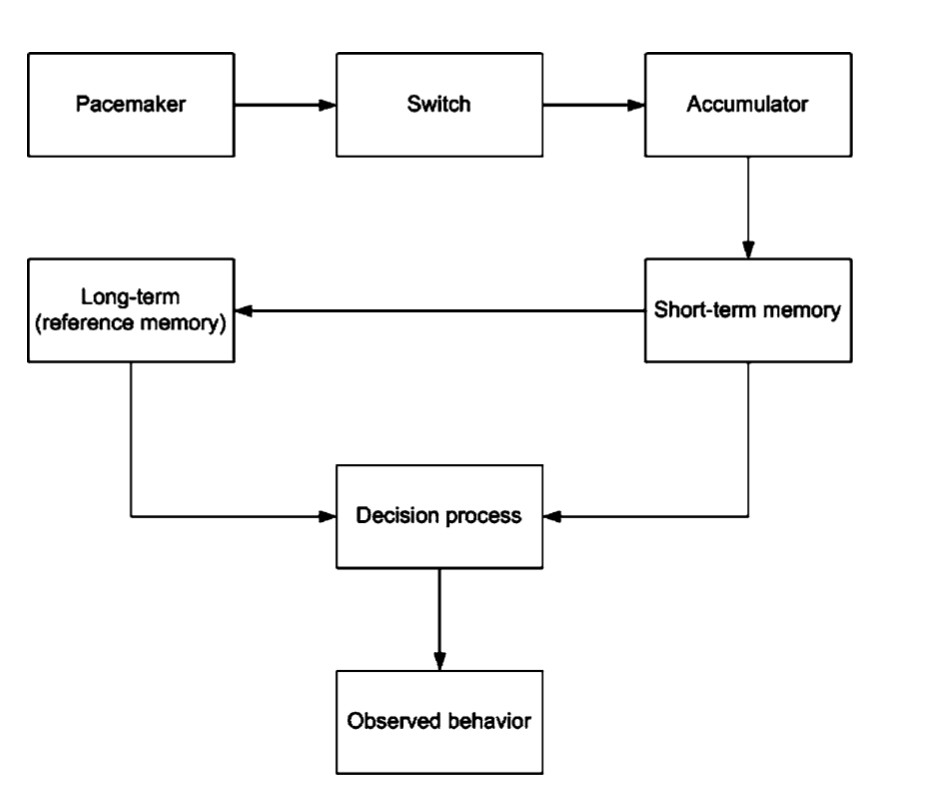
The prospective model says that an observer judges the duration of an
interval that is presently being experienced. This model assumes of an
"internal clock" with a pacemaker producing a sequence of time units
that are fed into an accumulator. Here the time units produced are
only registered when attention is directed to time. This makes
prospective model dual in nature as the observer has to switch focus
between temporal and non-temporal processes while the accumulator
collects the time units generated by the pacemaker in the background.
This model is also called the
Scalar Expectancy Theory. An internal
clock, working memory, a reference memory, and a decision process is
required as per this model. The onset of a stimulus causes the switch
to connecting the pacemaker and the accumulator to close allowing the
pulses or the ticks of the inner clock to flow. The offset of the
stimulus causes the switch to open, cutting the connection. The
resulting pulses represent the elapsed time and is sent to the working
memory. While timing an event the value in the working memory i.e. the
current time is compared to the value in the long term reference
memory i.e. the expected time. When the two values are close an action
is initiated; when they are far apart the action is stopped. To make
the comparison, the ratio of the two values are computed; when the
ratio is less than a certain value an action is initiated else no
action is initiated. The key point here is that a ratio of the two
values is compared instead of subtracting the absolute values i.e.
timing precision is relative to the size of the interval being timed.
Hence the name "Scalar". For eg. when timing a 10 second interval the
observed value might be precise upto 1 sec but if timing a 100 second
interval the precision might be about 10 seconds. However, a cyclical
pattern can be observed here. You need a reference memory to compare
the durations and these references are created from previous duration
experiences which themselves require duration judgments with
references that do not exist yet. How do you compare your first
duration when you have no stored pulse counts? The model works well
for explaining timing behaviour but struggles with the foundation
question of how temporal quality emerges initially. Clocks, were
developed on the original capacity we have to subjectively experience
time; they are a product of our mental, physical, and psychological
capabilities. We created the devices like clocks, chronometers,
watches, etc. because we could already perceive duration, sequence,
and a temporal flow. Clocks produce a measurable time for us which are
a result of having a temporal experience and we developed them to
explain the same experience; it is circular to explain how time works
by pointing to a clock.
The retrospective model of temporal processing says, the observer has
to estimate the already elapsed time interval duration. The observer
has to retrospectively estimate a given duration from the amount of
processed and stored memory contents i.e the duration has to be
constructed from memory. This model explains why certain events feel
longer in duration than others. Our experience of time is strongly
influenced by cognitive factors such as the amount of information
processed during the interval, the complexity of the stimuli, the way
the stimuli was stored in memory, memory load, level of attention,
etc. That is to say the more changing experiences we have during a
certain timespan the longer the duration is subjectively experienced.
So a routine activity if compared to a new or novel activity leads to
the perception of shorter duration. The judgments that we make in
retrospective about the duration are based on intervals spanning a few
seconds (short term memory) to a whole life time (long term memory). A
storage size metaphor is applicable here to explain this model.
Duration as a mental construction from memory is formed from the size
of the information stored in a given interval. It is the information
in the storage that determines duration experience. However, just like
the prospective model this model too does not account for all the
other modes of experiencing time. In fact it was shown through
experiments that if observers are asked to prospectively judge a time
interval and perform a complex task concurrently during the interval,
their perception of the interval duration shortens instead of
extending as a function of the amount of non durational information
processed in the interval. Another factor that strongly influences
retrospective duration estimate is contextual changes in the form of
environmental, psychological, interoceptive, etc. that makes the
observer think that the judged interval was longer in duration which
the storage size model cannot account for. The reliance of the
retrospective model on the human memory leads to flaws in itself.
Human memory does not work like computer memory. Information stored as
memory in the brain is constantly being shuffled, reinterpreted, and
interconnected with other memories; the information does not stay
static. The retrospective model assumes we have discrete information
units stored in our brains like computer files and we use them to
judge durations. When you recall something from your memory you are
not just recollecting the memory, you are also reconstructing the
experience.
As you can see, no matter what model we come up with to explain the
temporal experience it always falls short to account for the full
experience of time and keeps us looking for more ways to understand
how exactly the brain processes time.
Neural model of time
Currently we have two opposing models how these different timing
stimuli are processed in the brain. The hypothesis of the first model
is that there is a common mechanism that processes temporal
information across all the behaviours. This hypothesis is supported by
classical psychophysical (a branch of psychology that studies relation
between physical stimuli and mental phenomena) observations eg.
perception and motor production share common timing mechanisms and by
brain lesion and functional imaging studies. On the other hand the
hypothesis from the opposing model says the temporal processing is
ubiquitous and localised depending on the behavioural context. It
suggests that time processing does not have a dedicated parts in the
brain but rather a property that emerges from the normal functioning
of whatever brain networks are involved in the task. This view is
supported by modeling, brain slice recordings, and new psychophysical
approaches. There is an emerging third view which suggests the
existence of partially distributed timing mechanism, integrated by
main core structures of the cortico-thalamic-basal ganglia circuit
(CBGT) and the areas of the brain that are selectively engaged in the
behavioural task and is supported via recent psychophysical studies
and functional imaging meta analysis.
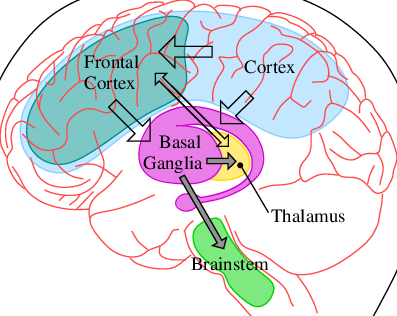
Studies done in retrospective model of timing have yielded the
hypotheses that the fronto striatal circuits loop between the
supplementary motor area (SMA), caudate putamen and pallidum which
form a part of basal ganglia are modulated by dopamine systems and are
critical for processing interval duration. Individuals with damage to
frontal lobes or injuries predominantly affecting the frontal areas
show impaired estimates of temporal intervals. In fact, various
neuroimaging studies show how dopaminergic systems affect our time
perception. Haloperidol is an antipsychotic medication that blocks
dopamine D2 receptors in the brain and it impairs the duration
discrimination abilities in healthy individuals. Studies in both
humans and animals have shown that dopamine agonists like
methamphetamine and antagonists like haloperidol influence the timing
process presumably by increasing and decreasing the clock speed
respectively. Individuals dependent on cocaine and methamphetamine who
have structural changes in the dopaminergic targets such as the
frontal cortex show impaired time processing on various timing tasks.
Hence, an intact dopamine neuro-transmission within the frontal areas
of the brain is a crucial requirement for accurate temporal processing
abilities.
Sensory Timing
All organisms extract temporal information from stimuli of all sensory
channels even if there is no time sensory organ. We are still not sure
how time is computed from the activation of different sensory systems
nor where is the temporal information extracted in this hierarchy of
sensory systems. But let's first understand the the general functional
arrangement of the auditory, visual and somatosensory (relating to a
sensation such as pressure, pain, or warmth which can happen anywhere
in the body) systems that are involved in the temporal processing
particularly in the hundreds of milliseconds range. These sensory
systems have the following commonalities: the sensory trigger of a
signal of the physical information into action potentials in the
sensory receptor; the projection of this information through the
thalamus to the primary sensory areas of the cerebral cortex; the
processing of the different aspects of the stimuli in the cortical
circuits of the brain; and finally the use of high order sensory
processing for perception, learning, memory and voluntary motor
action. Initially, time information could be extracted from the
trigger of the signal of the sensory input in the first relays of the
sensory systems involved. Not just that various studies show that
auditory modality has the strongest ability to extract temporal
information in the range of tens of milliseconds across the first
relays of the sensory processing which tells us that time is a
fundamental behavioural parameter for audition.
For vision, duration tuned cells show up in the first nodes of the
visual pathway. Some of the neurons respond best to stimuli that last
a specific amount of time. The time range is between 50 to 400
milliseconds. Interestingly, no such tuned cells were found in thr
thalamus for visual processing which suggests that time processing in
visual stimuli is localised in the first relays of neurons. However,
research has shown that temporal processing observed psychophysical
tasks in humans is more precise and accurate when the intervals are
defined by auditory than visual or tactile stimuli.
Another important property of temporal processing is that the
perceived time is affected by whether the intervals in the stimuli are
empty or filled. For auditory input empty stimuli could look like:
click - silence - click. For visual stimuli it could look like flicker
- darkness - flicker. The filled stimuli for auditory stimulus could
be a stream of noise or tone and for visual it could be just a flash
for the whole interval. Studies have shown that intervals with filled
stimuli feel longer than the stimuli with empty intervals. No single
mechanism fully explains the effect, it is the result of fundamental
differences in how the brain processes temporal information when
additional sensory processing is required versus when timing can
operate in isolation.
The temporal processing hence seems to be dependent on a specialised
group of cells in the early nodes of the sensory processing that are
tuned to the duration of auditory or visual stimuli. Also, the
multimodal temporal processing of tens of hundreds of milliseconds is
anatomy dependent (eyes vs ears) and audition clearly has an advantage
in the perception of passage of time.
Time Perception
The integration of duration information across all the sense seems to be dependent on the exstratriate cortex of the brain which is the region next to the occipital cortex which processes visual sensory information. The recognition and perception of passage of time across the senses can be used for planning a motor action, discriminating between intervals of time, categorisation tasks, etc. Multiple studies have been do to see how the brain performs interval learning and discrimination on multiple sensory modalities like audition and somatosensory stimuli. Subjects were trained to recognise a 70-200ms interval between stimuli and it was observed that intensive learning can generalise across modalities, skin location, and brain hemisphere. In one of the studies the subjects were trained on a base interval of 75ms and a target interval of 125ms and the discrimination of the intervals improved significantly on training of the two intervals. In fact interval training for somatosensory stimuli on one hand was generalised for the same interval on the other untrained hand. For auditory stimuli the learning transfer was dependent on improved temporal processing and not on more efficient memory or decision process. These findings not only support the view of a centralised or a partially overlapping distributed timing mechanism but also the existence of duration specific circuitry for temporal processing. Because of the learning the timing signals from the early cortical areas of the brain are sent to the core timing network of the brain which improves the efficiency of processing temporal information. fMRI studies show an early and late visual and auditory areas are activated during the perception of interval tasks. Overall, we see the idea emerging that temporal processing is a complex phenomena which involves cortical and subcortical structures of the brain.
Motor Timing
Time interval processing in the range of milliseconds is required for
various complex activities like speech perception and production,
perception and execution of music and dance, performance of sports,
etc. Time in music comes in different sequences and of different
durations; regular interval of different durations on a pulse forms a
meter. The ability to interpret the beat and rhythm helps us
appreciate music and be able to dance to it. Music and dance are
complex behaviour dependent on processing intricate loops of timing
information from the sensory organs or generating internally timed
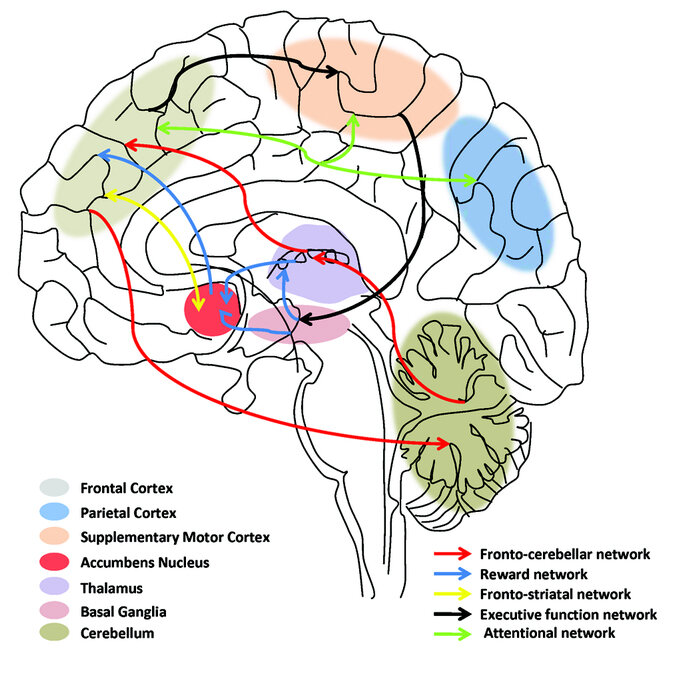
motor sequences in case of dancing. Various fMRI studies have shown
activation in cortico-thalamic-basal ganglia circuit when involved in
time perception activities which are the same areas that get activated
in planning and executing motor activities. This region of the brain
also includes medial premotor areas (premotor cortex) and
presupplementary motor areas (motor cortex). These studies provide an
evidence for CBGT being a core element of the timing network for
interval processing from the provided stimuli however, there seems to
be no evidence for its involvement in capturing the temporal
resolution during temporal processing.
Studies show the involvement of medial premotor areas and the
neostriatum (a cluster of nuclei in subcortical basal ganglia) for
duration tuning on a set of tapping tasks done with monkeys. Such
studies again confirm the existence of interval tuning in the core
timing network of the brain. Interval tuning here means that there are
a set of neurons in the CBGT tuned to a specific interval (let's say
100ms) where they fire with the highest intensity. For different
tapping intervals and modalities (visual cue, auditory cue) the
neurons showed an affinity for their tuned interval across all the
tasks. For instance a neuron tuned to 200ms stayed tuned to 200ms
whether the monkey was
- tapping to visual cues
- tapping to auditory cues
- tapping internally without a cue
- making single or multiple taps
This confirms the existence of tuned neurons which are context
independent for temporal processing in the CBGT. It's like having an
universal stopwatch for 200ms detection. In contrast, the neurons in
the lower level sensory processing (auditory and visual) are modality
specific, which means visual neurons only respond to visual stimuli
and auditory neurons only respond to auditory stimuli and hence
localised temporal resolution and processing.
In various experimental psychology studies it was observed that the
temporal processing improved significantly when there are repetitions
of the standard interval presented to the subjects.
Just Noticeable Difference is the amount
some aspect of a stimulus must be changed in order for a difference to
be noticeable, detectable at least half the time between two stimuli.
The standard interval is the interval chosen for various tasks in the
studies mentioned above. Multiple exposure to the standard interval
improved the accuracy of the subjects whether they were performing
visual or auditory tasks for time interval perception or temporalised
tapping. Recordings of the cells in premotor areas of monkeys
producing multiple intervals in a sequence showed temporal improvement
possibly due to an increase in the perceived or executed intervals.
The interval tunes cells in the CBGT showed a multiplicative scaling
i.e a proportional increase in their response for more produced
intervals. The decrease in the variance of the temporal information
could be because of the discharge rate of the interval tuned cells.
The discharge rate is the rate at which the neurons fire and is
measured in spiker per seconds or hertz (Hz). Higher the discharge
rate higher the neuron signal which leads to better noise to signal
ratio improving the overall accuracy.
One theory of why the neural circuitry of timing shares so much with
motor function is that our general sense of time has been developed
through actions since childhood. Developmental studies have shown that
young kids appear to represent time in motor terms; for them time is
how long does it take to do something. They are able to accurately
determine an interval if that interval has a motor action rather than
being empty. They also find it difficult to dissociate a duration from
the motor act itself. A young kid might think a task that required
more physical effort took longer even if the durations were identical.
It is possible that in case of temporal processing, the learned
associations between particular actions and their durations have been
engrained in the motor networks. Timing is crucial to perform any
motor activity. What is the point of swinging your bat at the wrong
time when the ball has already hit the stumps?
The human ability to perceive and generate precise timed intervals is most evident in musical perception and performance. Studies of brain potentials have indicated that human brain has this innate ability to predict rhythmic sensory events and error signals emerge when the music fails to mee the metric expectations. These error signals are measurable electrical patterns in the brain that indicate a mismatch between what we predicted would happen and what actually happened. Our sense of musical time is not passive; we are actively monitoring and predicting the temporal structure of music. It is evident that appreciating music does not just activate auditory cortex but it also engages our motor cortex to follow the rhythmic pattern engrained in the music. The joy of music comes from this ability to predict such rhythmic patterns. As we have seen the ability to predict sensory events and generate precisely timed actions seems not to only depend on the localised general purpose timing circuit, rather every neural network engaged in the sensory perception incorporate time as an essential feature of the information it is processing.